

Scientific publications list
Albicidin is extremely potent bactericidal antibiotic active against problematic Gram-negative bacteria however its mechanism of action was not understood at the molecular level. Now, a collaborative research team of Polish, German and British scientists used cryo-electron microscopy facility at SOLARIS to determine structure of albicidin bound to the Escherichia coli DNA gyrase complexed with 217 double-stranded DNA fragment.
Peptide toxin albicidin is produced by the bacterial plant pathogen Xanthomonas albilineans, which causes the devastating leaf scald disease in sugar cane. Albicidin is used by the pathogen to destroy plant chloroplasts, enabling its spread. Albicidin stops DNA replication in chloroplasts by targeting essential DNA topoisomerase DNA gyrase that is also present in all bacteria. Therefore, albicidin is extremely potent bactericidal antibiotic active against problematic Gram-negative bacteria however its mechanism of action was not understood at the molecular level. Now, a collaborative research team of Polish, German and British scientists used cryo-electron microscopy facility at SOLARIS to determine structure of albicidin bound to the Escherichia coli DNA gyrase complexed with 217 double-stranded DNA fragment.
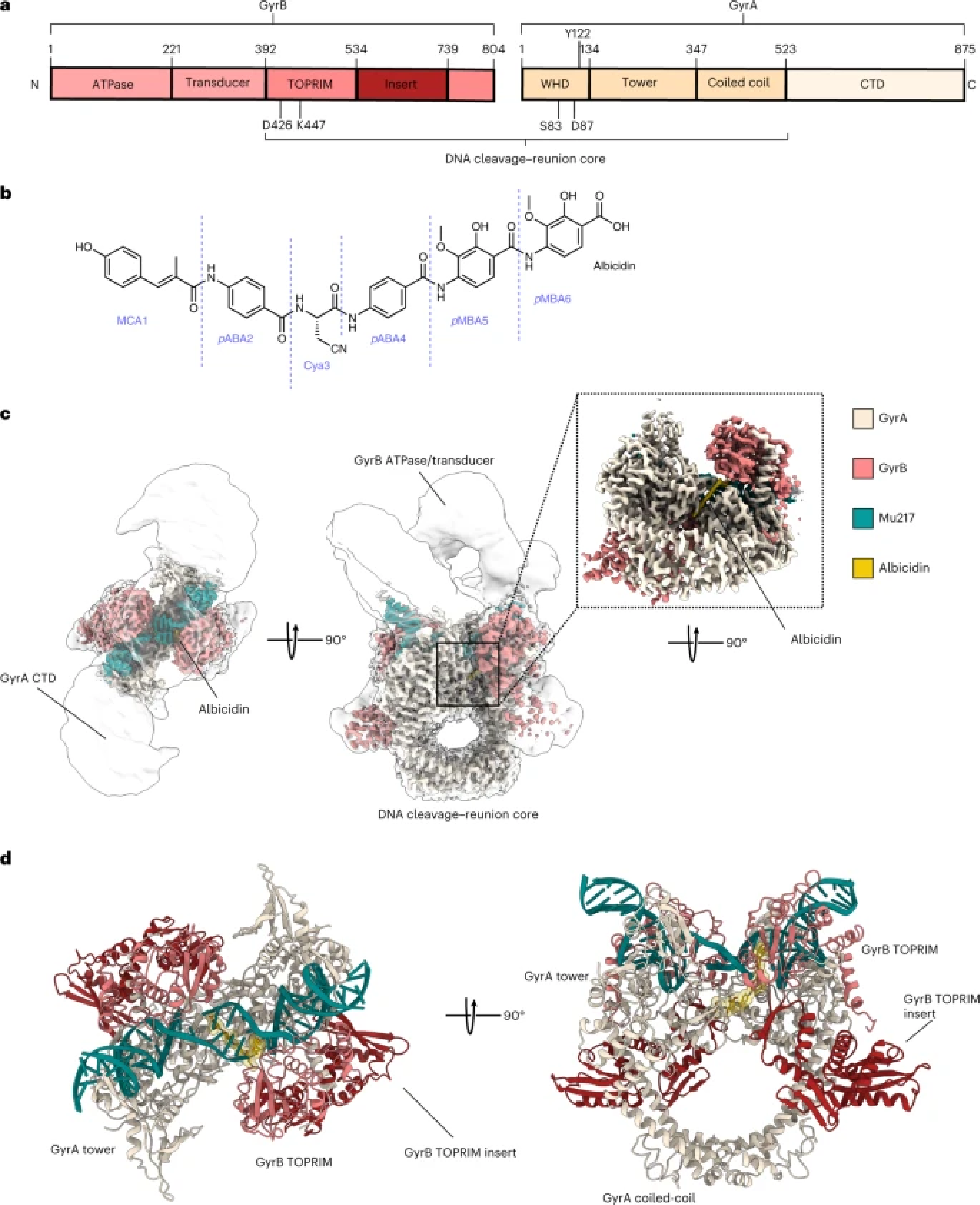
Figure 1. a, Scheme of GyrA and GyrB domains. The DNA cleavage–reunion complex is shown, along with the catalytic residue (Tyr122) and residues involved in quinolone resistance (GyrB Lys447 and Asp426 and GyrA Ser83 and Asp87). A consistent colour code is used throughout the manuscript: beige, GyrA; coral, GyrB. b, Chemical structure of albicidin. c, An overview of the Gyr–Mu217–albicidin cryo-EM map depicted as an overlay of two different contour level maps. Low-resolution contour (white) illustrates the position of GyrA CTDs and GyrB ATPase domains. High-resolution core part, including albicidin (in the zoomed-in image), is coloured according to the scheme in a: coral, GyrB; beige, GyrA; teal, DNA; yellow, albicidin. d, Cartoon representation of the overall model.
Development:
High-resolution (2.6 Å) structure of Gyr-Mu217-albicidin complex revealed a complex composed of two GyrA and two GyrB proteins, with 217 DNA wrapped around and cleaved, forming covalent bonds with catalytic tyrosine sidechains of the enzyme. Albicidin was clearly visible in the density: it uses a unique bicentric mode of binding, where N-terminal part of the molecule intercalated into DNA cleaved by the enzyme while C-terminal part wedges between two opposite GyrA monomers, jamming their movement and preventing progression of the catalytic cycle. We have carried out a structure-activity study and additionally determined structures of the improved synthetic albicidin derivatives that showed higher activity and better solubility and were active in animal models. These structures shown that the N-terminus of the molecule can be substituted by the flat aromatic moieties with known affinity to DNA such as quinoline or naphthalene. At the same time, albicidins were not at all affected by the gyrase mutations responsible for the fluoroquinolone resistance and therefore represent an entirely novel group of topoisomerase poisons and exciting drug candidate molecules. The structural understanding will now allow to further exploit a novel binding pocket and make more modifications to albicidin to improve its efficacy and pharmacological properties.
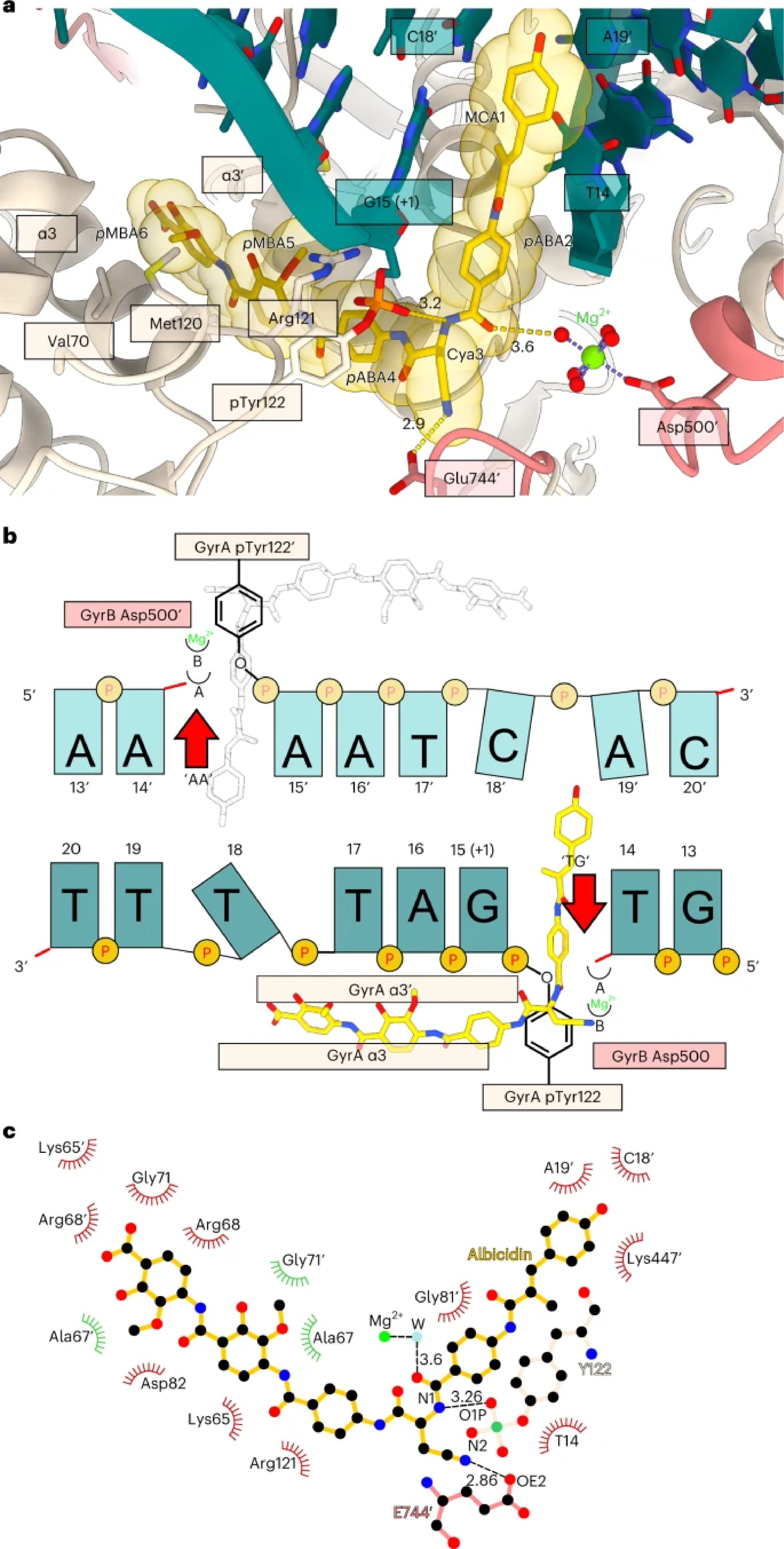
Figure 2. a, Enlarged view of the albicidin-binding site in the Gyr–Mu217–albicidin structure. Gyrase is represented as a cartoon, and albicidin as a stick representation. Van der Waals radii for albicidin atoms are shown as transparent yellow spheres. Two opposing GyrA helices α3 and α3′ at the dimer interface (DNA gate) form one part of the binding pocket, while DNA bases form another part. Distances (Å) between the modelled metal ion water shell and GyrB Glu744′ to Cya3 of albicidin are indicated. b, Schematic of albicidin binding in the context of the Mu cleavage site. Two potential binding pockets next to the scission sites ‘TG’ and ‘AA’ are labelled by the red arrows. Albicidin position is depicted by the sticks model (yellow), with the grey image indicating the potential alternative orientation not observed in the Gyr–Mu217–albicidin data. Two metal-binding sites, A and B, are indicated as half-circles. c, A LigPlot76 two-dimensional diagram of the albicidin-binding site. Hydrogen bonds and lengths (<4 Å) are indicated with dashed lines and the non-bonding and hydrophobic interactions (<4 Å) are labelled by the red and green spiked arcs, respectively. W, water coordinated to the metal (presumed Mg2+) ion.
Written by Ghilarov Dmitry
The publication can be found here: