
Kriomikroskopia SOLARIS (Cryo-EM Facility) oferuje przygotowanie próbek do badań kriomikroskopowych (witryfikacja), wstępną charakterystykę próbek (ang. sample screening) oraz zbieranie zestawów danych w ramach dostępnych technik badawczych: analizy pojedynczych cząstek (SPA), kriotomografii elektronowej (cET) i dyfrakcji elektronowej na mikrokryształach (MicroED).
Dodatkowo wspieramy naukowców i użytkowników przemysłowych w analizie danych eksperymentalnych zebranych w naszej pracowni.
Opis dostępnych technik badawczych kriomikroskopii elektronowej (Thermo Fisher Scientific)
- analiza pojedynczych cząstek (SPA) dostępna jest po kliknięciu w aktywny link.
- kriotomografia elektronowa (cET) dostępna jest po kliknięciu w aktywny link.
- dyfrakcja elektronowa na mikrokryształach (MicroED) dostępna jest po kliknięciu w aktywny link.
Przed i po eksperymencie – ważne informacje
- Aby uniknąć ewentualnych opóźnień eksperymentu i w pełni wykorzystać przyznany czas badawczy na kriomikroskopie Krios G3i, użytkownicy proszeni są o przekazanie opisanych próbek gotowych do pomiarów (zamrożonych na siatkach TEM i wstępnie scharakteryzowanych) na co najmniej tydzień przed zaplanowaną sesją pomiarową.
- Przeszkoleni użytkownicy mogą samodzielnie prowadzić badania wstępne przy użyciu mikroskopu Glacios pod nadzorem opiekuna infrastruktury.
- Zebrane dane eksperymentalne przechowywane będą przez 30 dni od zakończenia eksperymentu. Po tym okresie mogą zostać usunięte bez uprzedniego powiadomienia użytkownika.
- Przebadane próbki zachowane zostaną na okres 6 miesięcy od zakończenia eksperymentu. Po tym czasie mogą zostać usunięte bez uprzedniego powiadomienia użytkownika.
- Oferujemy konsultacje i wsparcie dla działań użytkowników w ramach eksperymentów kriomiorskopowych.
- Kriomikroskop Glacios (200 kV): 1 zmiana (shift) trwa 24 godz., z czego 8 godzin przeznaczamy na badania wstępne, a 16 godzin na zbieranie zestawów danych. Detektor Falcon 4 umożliwia rejestrację ~450 filmów/godz., co przekłada się na blisko 7000 filmów w ciągu zmiany (ok. 4,2 TB danych).
- Kriomikroskop Krios G3i (300 kV): 1 zmiana (shift) trwa 60 godz., z czego 55 godzin przeznaczamy na zbieranie danych. Detektor K3 umożliwia rejestrację ~680 filmów/godz., co przekłada się na blisko 40000 filmów w ciągu zmiany (~16 TB danych).
- Aby usprawnić proces kopiowania danych zalecamy dostarczenie przenośnego dysku 3,5” (ze stacją dokującą) w celu kopiowania danych „w locie”. Możliwe jest także skopiowanie danych użytkownika na zewnętrzne serwery danych.
- Analizę zebranych danych można przeprowadzić korzystając z superkomputera Athena zlokalizowanego w ACK Cyfronet AGH.
Metodologia
Metodologia prac badawczych w technice kriomikroskopii elektronowej obejmuje 5 etapów przedstawionych na poniższym schemacie (rys. 1.). Cały proces rozpoczyna preparatyka i oczyszczanie próbki ze wstępną charakterystyką biochemiczną. Następnie próbka poddawana jest witryfikacji na siatkach mikroskopowych oraz badaniom wstępnym na kriomikroskopie w celu oceny jej jakości, grubości lodu oraz dystrybucji cząsteczek w nim zawartych. Dla optymalnych próbek, zbierany jest właściwy zestaw danych, służący rozwiązaniu struktury przestrzennej badanej próbki.

Rys.1. Od biochemii do struktury przestrzennej makromolekuł – kolejne kroki badawcze w technice kriomikroskopii elektronowej.
Preparatyka materiału biologicznego
Dla powodzenia badań kriomikroskopowych i określenia struktury przestrzennej badanej cząsteczki kluczowe znaczenie ma jakość próbki. Próbki do badań kriomikroskopowych powinny charakteryzować się wysoką czystością, stabilnością oraz homogennością konformacyjną. Należy również zwrócić uwagę na odpowiedni dobór buforu stabilizującego próbkę, bez zbędnych dodatków, które mogą wpływać na kontrast obrazu (np. glicerolu).
Preparatykę próbki można przeprowadzić korzystając ze standardowych metod oczyszczania białek rekombinowanych, takich jak np. różnego rodzaju chromatografie (powinowactwa, jonowymienna) oraz sączenie molekularne (ang. size exclusion chromatography, SEC). Czystość preparatu można ocenić przy pomocy elektroforezy w żelu poliakryloamidowym (SDS-PAGE), a homogenność próbki przy pomocy układu SEC–MALS (wielokątowe rozpraszanie światła, ang. multi angle light scattering) lub dynamicznego rozpraszania światła (DLS).
Przykładowa analiza biochemiczna wyizolowanych i oczyszczonych białek została przedstawiona na rys. 2. Dalsze analizy, w tym aktywność enzymatyczna, spektroskopia optyczna i analiza elektroforetyczna wykazały, że badana frakcja próbki zawierała pełny zestaw podjednostek Cyt b6f i była aktywna enzymatycznie, stąd została rekomendowana do badań kriomikroskopowych. (Sarewicz et al., doi:10.1126/sciadv.add9688).
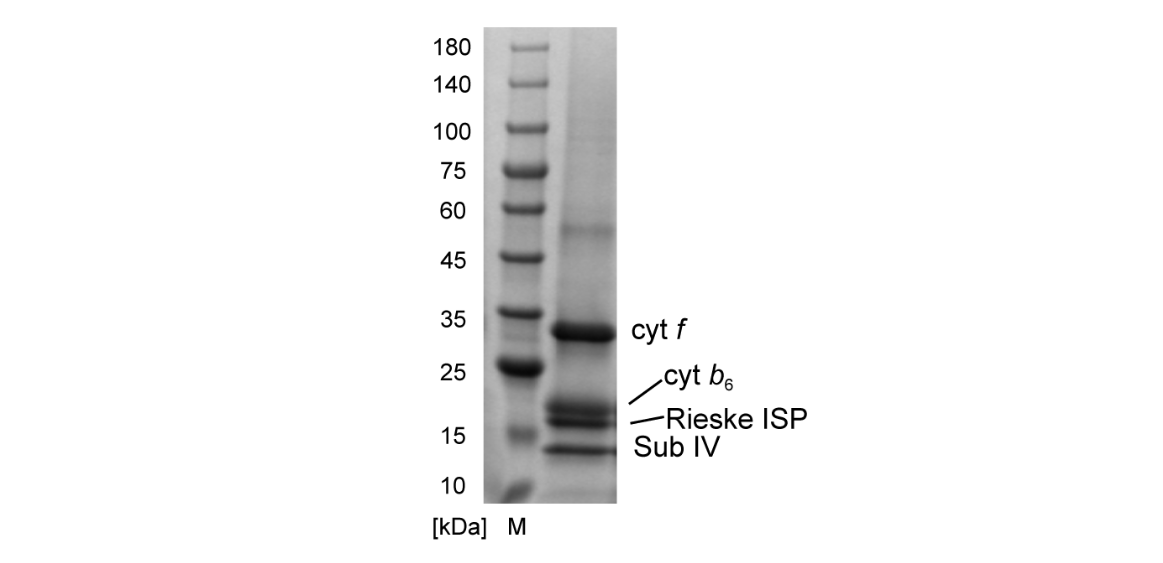
Rys. 2. Żel SDS–PAGE obrazujący czystość próbki kompleksu Cyt b6f zobrazowanego w kriomikroskopie. Żel wybarwiono barwnikiem Coomassie Briliant Blue. Każdemu prążkowi odpowiada białko o określonej masie molekularnej. Sarewicz et al., doi:10.1126/sciadv.add9688
Preparatyka próbek kriomikroskopowych Cryo-EM
- napylarka próżniowa (Safematic CCU-010 Compact Coating Unit) - urządzenie to wykorzystuje się do przygotowania powierzchni komercyjnych siatek mikroskopowych poprzez ich aktywację w niskoenergetycznej plazmie, tworzonej przez wyładowanie jarzeniowe w rozrzedzonym gazie. Zwiększona hydrofilowość folii węglowej (polimerowej) siatki TEM poprawia zwilżalność jej powierzchni, a co za tym idzie pomaga zoptymalizować rozkład cząsteczek w lodzie. Oddziaływaniem plazmy na powierzchnię siatek można sterować poprzez odpowiednie ustawienie: wysokości stolika z siatkami względem katody, prądu wyładowania i czasu ekspozycji. Napylarkę można również wykorzystać do napylania nanometrycznych warstw amorficznego węgla.
- robot do witryfikacji próbek kriomikroskopowych (Vitrobot Mark IV Thermo Fisher Scientific). Kropla buforu z próbką umieszczana jest na siatce mikroskopowej, jej nadmiar jest odciągany przez papier filtracyjny, po czym preparat zostaje z dużą szybkością, zanurzony w ciekłym etanie, co prowadzi do uwięzienia cząsteczek próbki w amorficznym lodzie. Proces mrożenia przebiega w komorze środowiskowej i jest sterowany poprzez graficzny interfejs użytkownika, co zapewnia większą jego powtarzalność. Procesem mrożenia można sterować poprzez odpowiednie ustawienie programu witryfikacji tj. dostosowania parametrów m.in.: względnej wilgotności komory (do 100%), temperatury komory (4-60 °C), czasu i siły odciągania buforu z próbką, a także czasu inkubacji próbki na siatce przed zamrożeniem.
Próbka
- zastosowanie siatek pokrytych ultracienką warstwą tlenku grafenu - wiele biomolekuł ma silne powinowactwo do węgla, przez co można polepszyć ich dyspersję czy dodatkowo je zagęścić w lodzie,
- dodatek detergentów w buforze - obniża napięcie powierzchniowe,
- dostosowanie metody zbierania danych, tak by obejmowała również nachylone ekspozycje próbek.
Siatki mikroskopowe
W zależności od potrzeb projektu użytkownika, optymalną siatkę można dobrać z szerokiej gamy komercyjnie dostępnych siatek mikroskopowych (typ materiału z którego wykonano siatki TEM, kształt i wielkość otworów siatki (tzw. mesh), typ pokrycia siatki (np. formvar, tlenek grafenu, węgiel amorficzny, …), rodzaj pokrycia siatki czy wielkości otworów folii nośnej (np. quantifoil, lacey, holy, continuous, …) itd.
Z technicznego punktu widzenia zbierania zestawów danych kriomikroskopowych oraz ich późniejszej analizy, folia wspierająca próbki powinna zapewniać zarówno wysoką stabilność dla próbki, jak i niski poziom sygnału tła. Ponadto geometria otworów w folii powinna być optymalna dla danej metody eksperymentalnej, tak aby uzyskać wysoką przepustowość eksperymentu i zebrać możliwie jak najwięcej danych w jak najkrótszym czasie.
Przykładowo dla techniki SPA białek i kompleksów białkowych optymalna byłaby siatka Cu 200 mesh z pokryciem typu quantifoil o geometri R2/1 czyli układu okrągłych otworów o średnicy 2 µm i odległości pomiędzy otworami równej 1µm. Niewiele mniejszą szybkość zbierania danych uzyskuje się też dla siatek R1.2/1.3. Na ogół próbka o większych wymiarach zwłaszcza w przypadku całych komórek czy bakterii, będzie wymagała większych rozmiarów otworów w pokryciu siatki (np. lacey carbon) i ewentualnie dodatkowego wsparcia (quantifoli, holey carbon), Tam gdzie stosuje się technikę kriotomografii elektronowej, czy przy badaniach próbek wymagających pochyłu siatki w trakcie zbierania danych, często dobiera się siatki o większych otworach (np. siatka Cu 100 mesh), co pozwala zachować stałe pole widzenia przy większych kątach pochyłu.
Transfer siatek z próbkami
Do przenoszenia zamrożonych preparatów na siatkach mikroskopowych używamy opatentowanych uchwytów tzw. AutoGrids i kompatybilnych narzędzi (Thermo Fisher Scientific). Zastosowane uchwyty umożliwiają wymianę próbek pomiędzy mikroskopami oraz ich bezpieczne przechowywanie (Rys. 3).

Rys. 3. Przygotowanie próbek do badań kriomikroskopowych: klipowanie siatki z próbkami użytkowników (a), przekładanie ich do kasety (b), a następnie umieszczanie kasety z próbkami w zautomatyzowanym podajniku (ang. autoloader) kriomikroskopu Krios G3i (c). Kadr z video: CiekaWizja Wiktor Niedzicki – Synchrotron SOLARIS.
Optymalizacja próbek
- jakość, grubość i jednorodność lodu na siatce mikroskopowej,
- stężenie, integralność, stabilność i dystrybucja cząsteczek próbki w lodzie (czy cząsteczki ulegają agregacji lub dekompozycji).
- 155x – jakościowy przegląd oczek siatki mikroskopowej pod kątem gradientu grubości lodu,
- 700x – wybranie oczka siatki z optymalną grubością lodu,
- 13 500x – jakościowa ocena lodu z cząsteczkami w otworach folii polimerowej,
- 190 000x – obrazowanie cząstek docelowych zawieszonych w lodzie.

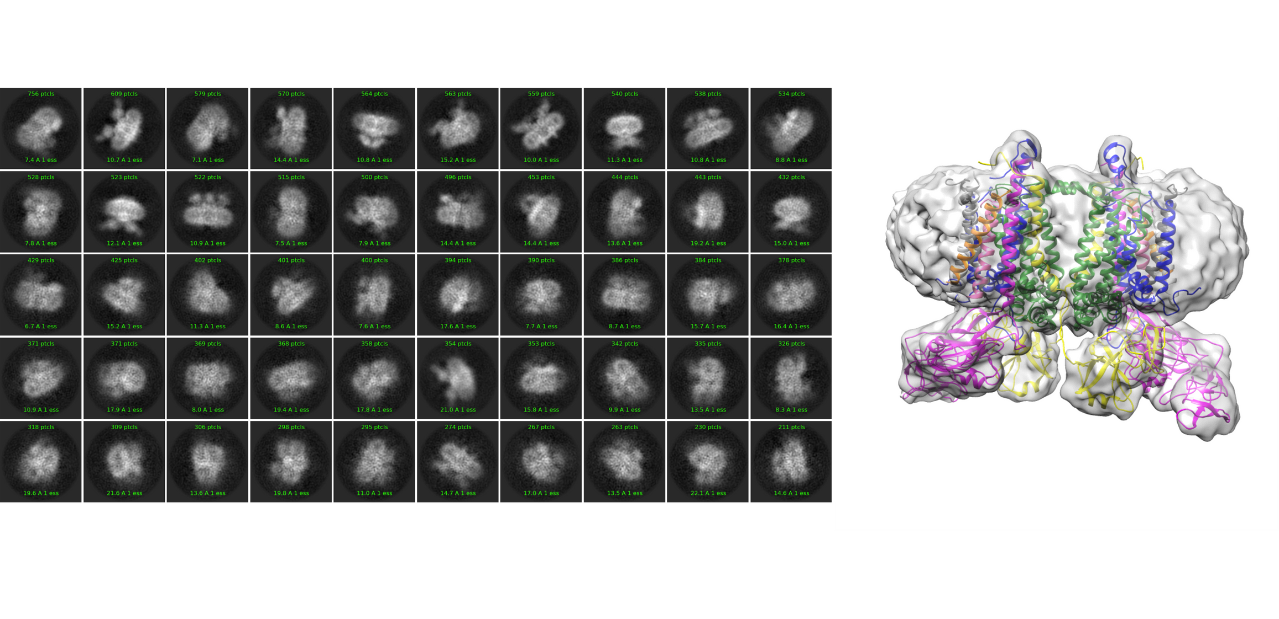
W wyniku analizy danych zarejestrowanych metodami cryo-EM uzyskuje się tzw. mapy potencjału lub mapy gęstości. Ten obiegowy skrót myślowy oznacza dane objętościowe zawierające zmierzony potencjał Coulombowski elektronów wiązki rozpraszanych na atomach próbki, jako reprezentację badanej cząsteczki. Interpretacja zwykle obejmuje inspekcję izopowierzchni zrekonstruowanej mapy za pomocą oprogramowania do wizualizacji, a następnie ręczne lub automatyczne opisywanie jej cech za pomocą współrzędnych atomowych i późniejsze udokładnianie modelu.
Z danych SPA rejestrowanych na mikroskopie Glacios użytkownicy rutynowo określają trójwymiarowe struktury makrocząsteczek w rozdzielczościach z przedziału 4-8 Å dla małych cząstek (<200 kDa) oraz poniżej 4 Å dla cząstek średnich i dużych (≥400 kDa).
Strategia zbierania danych przy użyciu kriomikroskopów
- Kriomikroskop Glacios (200 kV): 1 zmiana (shift) trwa 24 godz., z czego 8 godzin przeznaczamy na badania wstępne, a 16 godzin na zbieranie zestawów danych. Detektor Falcon 4 umożliwia rejestrację ~450 filmów/godz., co przekłada się na blisko 7000 filmów w ciągu zmiany (ok. 4,2 TB danych). Z danych SPA rejestrowanych na mikroskopie Glacios użytkownicy rutynowo określają struktury przestrzenne makrocząsteczek w rozdzielczościach z przedziału 4-8 Å dla cząstek małych (<200 kDa) oraz poniżej 4 Å dla cząstek średnich i dużych (≥400 kDa).
- Kriomikroskop Krios G3i (300 kV): 1 zmiana (shift) trwa 60 godz., z czego 55 godzin przeznaczamy na zbieranie danych. Detektor K3 umożliwia rejestrację ~680 filmów/godz., co przekłada się na blisko 40000 filmów w ciągu zmiany (~16 TB danych). Z danych SPA rejestrowanych na mikroskopie Krios G3i użytkownicy rutynowo określają struktury przestrzenne makrocząsteczek w rozdzielczościach z przedziału 2-4 Å (aktualny rekord 1,93 Å).
| Analiza Pojedynczych Cząstek (SPA) | ||
|---|---|---|
| Mikroskop | Krios G3i | Glacios |
| Napięcie przyspieszające | 300 kV | 200 kV |
| Całkowita dawka mierzona bez próbki | 40 e/A2 | 40 e/A2 |
| Rozmiar piksela [A/px] | 0.846 A/px | 0.73 A/px |
| Aberracja sferyczna | 2.7 mm | 2.7 mm |
| Liczba ramek na film | 40 | 40 or EER fractions |
| Typ danych | TIFF/MRC | EER/MRC |
| Powiększenie | 105 000x | 190 000x |
| Wartości rozogniskowania | -1.8, -1.5, -1.2, -0.9, -0.6 µm | -2.0, -1.5, -1.0 µm |
| Ekspozycje na oczko folii | 6-8 (/2µm) | 1-4 (/2µm) |
| Filtr energii | Gatan BioQuantum-slit 20eV | N/A |
| Detektor | Gatan K3 | TFS Falcon 4 |
| Tryb pracy kamery | counting mode | counting mode |
| Szybkość zliczeń detektora | 15-16 e/px/s | 6-8 e/px/s |
| Rozdzielczość obrazu | 5760 x 4092 | 4096 x 4096 |
| Typ filmu | .tiff gain corrected | .eer gain not corrected |
| Kriotomografia elektronowa (cET) | ||
|---|---|---|
| Mikroskop | Krios G3i | Glacios |
| Napięcie przyspieszające | 300 kV | 200 kV |
| Aberracja sferyczna | 2.7 mm | 2.7 mm |
| Rozmiar piksela .mrc | 1.69 A/px | 4.00 A/px |
| Rozmiar piksela .tiff | 0.846 A/px | - |
| Rozmiar piksela .eer | N/A | 2.00 A/px |
| Całkowita dawka mierzona bez próbki | 100-150 e/A2 | 100-150 e/A2 |
| Liczba ramek na film | 10 | frakcje EER |
| Strategia zbierania danych | dose symmetric | dose symmetric |
| Zakres serii kątowej (51 filmów) | -50 to +50 deg, step = 2 deg | -50 to +50 deg, step = 2 deg |
| Stałe rozogniskowanie | -5 um | -7 um |
| Powiększenie | 53 000x | 36 000x |
| Apertura C2 | 50 um | 150 um |
| Detektor | Gatan K3 | TFS Falcon 4 |
| Tryb pracy kamery | counting super resolution | counting mode |
| Szybkość zliczeń detektora | 15-16 e/px/s | 6-8 e/px/s |
| Rozdzielczość obrazu | 5760 x 4092 | 4096 x 4096 |
| Typ filmu | .tiff gain corrected | .eer gain not corrected |
| Filtr energii | Gatan BioQuantum - slit 40eV | N/A |
Rekonstrukcja i wizualizacja struktury w 3D
- Analizę zebranych zestawów danych można przeprowadzić korzystając z zasobów superkomputerów Cyfronetu (Athena).
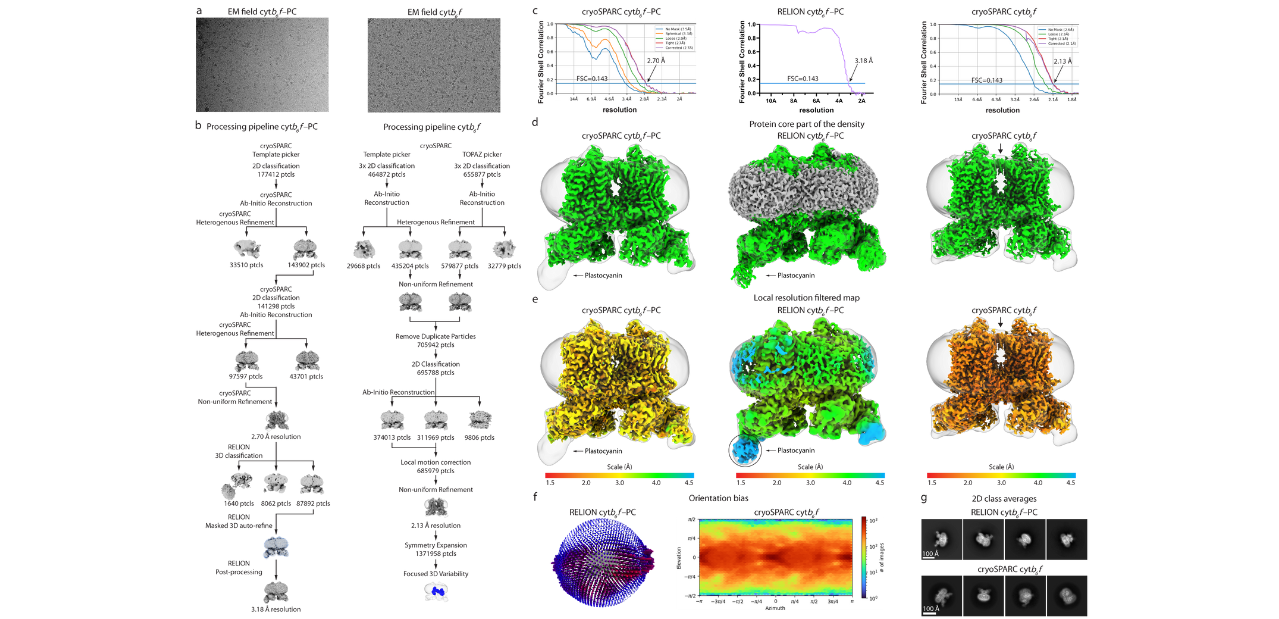
Rys. 7. Metodologia przetwarzania zestawów danych kriomikroskopowych dla kompleksów Cyt b6f – PC i Cyt b6f. (a) Reprezentatywne obrazy kriomikroskopowe, (b) kolejne kroki przetwarzania zestawu danych Cryo_EM, (c) korelacja powłok Fouriera (FSC). Niebieskie linie wskazują FSC=0,143 z zaznaczonymi odcięciami rozdzielczości rekonstrukcji, (d) gęstość rdzenia białkowego zaznaczona kolorem zielonym (wyostrzona mapa) oraz nałożona z półprzeźroczystą gęstością odpowiadającą powłoce lipidowej (wyostrzona mapa wygładzona gaussowsko). Wszystkie mapy konturowane do poziomu RMS 5, (e) zakres rozdzielczości wszystkich map na poziomie RMS 5, z plastocyjaniną konturowaną do poziomu RMS 3 dla mapy z RELIONa, (f) rozkład kątowy Eulera cząstek Cyt b6f – PC dający wkład do ostatecznej rekonstrukcji w RELIONie i rozkład kątowy cząstek obliczony w CryoSPARCu dla mapy Cyt b6f. Mapa temperaturowa przedstawia liczbę cząstek dla każdego kąta obserwacji, (g) wybrane uśrednione klasy 2D cząstek wchodzących do rekonstrukcji map Cyt b6f – PC i Cyt b6f w RELIONie. (Sarewicz et al., doi:10.1126/sciadv.add9688)
Infrastruktura IT: przechowywanie, transfer i procesowanie danych pomiarowych
- Obecnie dostępna przestrzeń dyskowa na serwerach SOLARIS to 231 TB dla danych z kriomikroskopu Krios G3i oraz 1,4 PB dla danych z mikroskopu Glacios. Przestrzeń dyskowa kriomikroskopu Glacios oparta jest na rozwiązaniu Dell PowerScale, zapewniającym bezpieczeństwo gromadzonych danych, przy jednoczesnej ergonomii administracji.
- Na potrzeby obliczeń i analizy danych kriomikroskopowych zakupiony został serwer obliczeniowy wyposażony w dwa CPU (łącznie 72 wątki), kartę graficzną Tesla V100 z 32 GB pamięci, co znacznie przyspiesza obliczenia. System wspomagany jest przez 750 GB pamięci RAM.
- Dla użytkowników dostępne są 3 stacje robocze GPU do wstępnego przetwarzania danych kriomikroskopowych.
- Transmisja danych pomiędzy podsystemami odbywa się po łączu o przepustowości 10 Gbps.
Przechowywanie próbek
- Aby uniknąć ewentualnych opóźnień eksperymentu i w pełni wykorzystać przyznany czas badawczy użytkownicy proszeni są o przekazanie próbek gotowych do pomiarów (zamrożonych na siatkach TEM oraz dobrze opisanych) na co najmniej tydzień przed zaplanowaną sesją kriomikroskopową.
- Użytkownicy mogą korzystać z systemów do magazynowania gotowych preparatów dostępnych w SOLARIS.
- Przebadane próbki są przechowywane do 6 miesięcy od daty zakończenia eksperymentu. Po tym okresie próbki są usuwane bez uprzedniego powiadomienia użytkownika.